- Applications
-
Products
-
Liquid Handling
- firefly Accelerate genomic research with innovative all-in-one, compact liquid handling
- mosquito Nanolitre liquid handling technology performs ‘traditional’ tasks at a fraction of the volume, and higher speeds
- dragonfly Delivers accurate and repeatable nanolitre to milliliter dispensing
- apricot Automated liquid handling instrumentation for convenient general use across your entire team
- Sample Preparation
-
Sample Management
- comPOUND A scalable, reliable, and secure compound management solution
- BioMicroLab Easy-to-use sample management automation instruments
- arktic Robust biospecimen storage and management down to -80°C
- lab2lab Novel sample and data transfer network system
- comPACT Reliable and efficient -20°C storage and retrieval has never been more accessible
-
Liquid Handling
-
About
- Company With a focus on liquid handling, sample preparation and sample management, our expert teams create state-of-the-art solutions that scientists and researchers can trust Culture We have one overarching mission: to work together to accelerate life science research. Through our innovative solutions and state-of-the-art tools, we believe we can make a real difference to human health Partners Collaboration is key in our mission to make a real difference to human health. Partnering with application leaders globally, we co-create to solve new challenges across the life sciences. Innovation From the initial prototype through to manufacturing, installation and beyond, we bring a problem-solving mindset and technical expertise to drive innovation
-
Executive Leadership
 Through strategic guidance, visionary thinking, and a relentless pursuit of excellence, our senior executives steer SPT Labtech towards achieving its mission of making a real difference to human health through solving advanced laboratory challenges.
Learn more
Through strategic guidance, visionary thinking, and a relentless pursuit of excellence, our senior executives steer SPT Labtech towards achieving its mission of making a real difference to human health through solving advanced laboratory challenges.
Learn more 
-
View all
Board of Directors
 Our Board of Directors are committed to driving the long-term success and sustainability of SPT Labtech, providing expert guidance and oversight to execute the company’s ambitious commercial strategy.
Learn more
Our Board of Directors are committed to driving the long-term success and sustainability of SPT Labtech, providing expert guidance and oversight to execute the company’s ambitious commercial strategy.
Learn more
-
Knowledge Base
- Resources Our wide range of insightful resources include videos, whitepapers, eBooks, application notes and more Events & Webinars Meet the SPT team at events all over the globe and virtually via our webinars Podcast We chat with innovators and leaders from across the community to gain their unique insights. News Latest news from SPT Labtech globally Blog Our latest blog posts feature trends in research, innovative techniques and new technology
-
09 July, 2025
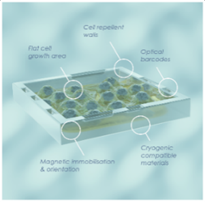 SPT Labtech and Semarion Collaborate to Advance Automated Cell-Based Assay Workflows
Continue reading
SPT Labtech and Semarion Collaborate to Advance Automated Cell-Based Assay Workflows
Continue reading
-
23 June, 2025
 SPT Labtech Named Illumina Qualified Methods Provider Following Successful Automation of Illumina DNA Prep on firefly® Platform
Continue reading
SPT Labtech Named Illumina Qualified Methods Provider Following Successful Automation of Illumina DNA Prep on firefly® Platform
Continue reading
-
23 April, 2025
 A trusted partner in drug discovery: Viral Patel’s 20-year journey with mosquito®
Continue reading
A trusted partner in drug discovery: Viral Patel’s 20-year journey with mosquito®
Continue reading
10
- Careers



- Home
- Resources
Resources
Explore insightful resources that include videos, whitepapers, eBooks, application notes and more
- 3D Cell Culture
- Assay Setup
- Automated Assay Development
- Biostorage and Biobanking
- Chemical Crystallization
- Compound Management
- CryoEM
- Crystal Screen Optimization
- DNA Normalization
- DNA Quantification
- Hanging Drop Crystallization
- High Throughput Screening
- Lipidic Cubic Phase (in meso)
- Magnetic Bead Clean-up
- Medicinal Chemistry
- NGS Library Prep
- PCR and qPCR
- Protein Crystallography
- Reformat
- Sample Tracking
- Single Cell Genomics
- Single Particle Analysis
- Sitting Drop Crystallization
- Synthetic Biology
- Transcriptomics
- Tube Cap/Decap
- Tube Labeling
Not sure which apricot liquid handler is right for you? Join Product Manager Jonathan Faherty as he breaks down the apricot S1, S3, PP5, and DC1—highlighting what each system does, who it’s best suited for, and how to choose the perfect fit for your lab's needs.
Read moreIn this session, Sara Talbot, Senior Research Scientist at AstraZeneca, shares her experience using the NEBNext UltraExpress RNA Library Prep Kit in combination with our firefly platform to prepare 96 RNA-seq libraries in a single day. This powerful integration of reagents and automation not only streamlined the library preparation process but also significantly reduced time at the bench and improved reproducibility across samples.
Read moreThe Illumina DNA Prep kit is an optimized, user-friendly workflow for preparing normalized and ready-to-sequence libraries in under four hours, from a wide range of DNA inputs. Illumina DNA Prep delivers reliable results - consistent insert sizes, uniform coverage, and optimized performance for a range of sample types and DNA input amounts1,2. Illumina DNA Prep relies on bead-based tagmentation technology where magnetic bead-based transposome complexes fragment DNA and ligate adapter sequences in a single 15-minute reaction. PCR is then used to add indexes to the DNA fragments and generates libraries compatible with all Illumina sequencing platforms. The amplified libraries are purified using Illumina Purification Beads. The overall process eliminates the need for library quantitation before library pooling and sequencing, minimizing bias and error, and resulting in highly reproducible sequencing data1,2. Here we show how the Illumina DNA Prep workflow, based on Illumina DNA Prep Reference Guide (1000000025416 v11), was automated on firefly®, in a two-part protocol with a single off-deck thermocycler step. Details of the firefly® workflow are described, and data is
Read moreAs sequencing costs continue to decrease, sample and library preparation have emerged as the critical bottleneck in modern genomics workflows. Core facilities face mounting pressure to process thousands of samples with accuracy and precision while maintaining competitive pricing and rapid turnaround times. In this session, you'll hear from Taylor Fennelly, a researcher at the Biopolymers Facility (BPF) at Harvard Medical School, who will share their success story implementing SPT Labtech’s firefly for high-throughput, high-quality library generation. Our very own firefly product manager, Denise Grovewood will demonstrate how this compact, all-in-one automated liquid handling solution is specifically designed to overcome the bottlenecks faced by sequencing cores.
Read moreThe Illumina DNA Prep kit is an optimized, user-friendly workflow for preparing normalized and ready-to-sequence libraries in under four hours, from a wide range of DNA inputs. Illumina DNA Prep delivers reliable results - consistent insert sizes, uniform coverage, and optimized performance for a range of sample types and DNA input amounts
Read moreThe application of AI in drug discovery relies on high-quality, reproducible training datasets. Traditional screening campaigns focus on identifying potent hits, but ML-driven drug discovery requires comprehensive potency evaluation across entire compound libraries. Here, we introduce a partial concentration-response curve (pCRC) approach that estimates potency using just two data points per compound. We onboarded a panel of 65 diverse kinases and screened 7000 compounds against the panel at ATP concentrations near Kₘ to minimize modality bias, achieving a mean robust Z’ of 0.74 across all targets. A direct comparison of 100 fragments tested in both 2-point pCRC and conventional 11-point CRC formats demonstrated excellent correlation, confirming that our pCRC methodology produces high-quality data suitable for ML model training. The integration of our automation platform, including SPT Labtech’s dragonfly® discovery, with automated data pipelines enabled the generation of 221,000 high-quality ML-ready data points per day, accelerating the development of foundation model training for drug discovery.
Read moreThis video gives a tutorial on how to dispense Matrigel and PeptiMatric hydrogels using the Dragonfly discovery for use cases such as for 3D Cell culture. The dragonfly® discovery is a low volume, positive displacement, non-contact dispenser. The instrument is available in three configurations that differ by the number of dispense heads (3, 6, 10) or channels. Channels can be combined for fast bulk reagent dispensing (as demonstrate3d in this video), or, used independently to dispense any volume of any liquid class into any well at any time.
Read moreThis technical note provides supporting information for the firefly protocols listed below. These protocols are based on the KAPA EvoPrep Kit Operators Manual v1.2 and are available to download from the firefly community. Here, we outline protocol run times, parts required and provide details on the steps performed in each protocol.
Read moreThe KAPA EvoPrep Kits are the latest high-performance, streamlined, and automation-friendly library preparation solutions from Roche. SPT Labtech firefly protocols have been developed to run the KAPA EvoPrep and KAPA EvoPrep PCR-Free library preparation workflows - using the 96-well plate format of the KAPA EvoPrep Kit and the KAPA Unique Dual-Indexed Adapter Kit.
Read moreThis technical note provides supporting information for automation of the NEBNext Ultra II FS DNA Library Prep kit on firefly. The firefly protocols are listed below and are available to download from the firefly community. Here, we outline protocol run times, parts required and provide details on the steps performed in each protocol.
Read moreThe NEBNext® Ultra™ II FS DNA Library Prep Kit for Illumina is optimized for the construction of high quality, high yield libraries using a broad range of DNA input amounts. The library preparation workflow uses NEBNext FS (Fragmentation System) enzymatic fragmentation reagents, eliminating the need to mechanically shear the DNA input. The workflow also uses a single enzyme mix to perform the fragmentation, end repair and dA-tailing, which greatly simplifies the workflow. Here, we demonstrate how this workflow has been automated on firefly. We provide details on the steps performed in each protocol, together with data to demonstrate the performance of these protocols in generating high quality libraries for sequencing.
Read moreIn vitro cell cultures are essential for modelling disease, but traditional 2D cell culture systems fall short in replicating the complex microenvironments of native tissues. This limitation hinders the translation of in vitro findings to in vivo contexts and contributes to the high attrition rates seen in drug discovery pipelines.
Read more